what does it mean to say that a mutation is deleterious?
Is it possible to have "besides many" mutations? What about "as well few"? While mutations are necessary for evolution, they can damage existing adaptations as well.
What is a mutation?

The diversity of beetle species.
Genetic mutation is the basis of species diversity among beetles, or any other organism.
© 2009 Courtesy of John C. Abbot, Abbott Nature Photography. All rights reserved. ![]()
Mutations are changes in the genetic sequence, and they are a main cause of diversity among organisms. These changes occur at many unlike levels, and they can take widely differing consequences. In biological systems that are capable of reproduction, nosotros must first focus on whether they are heritable; specifically, some mutations affect merely the individual that carries them, while others affect all of the carrier organism's offspring, and further descendants. For mutations to affect an organism'south descendants, they must: 1) occur in cells that produce the adjacent generation, and ii) affect the hereditary material. Ultimately, the interplay between inherited mutations and ecology pressures generates diverseness amid species.
Although various types of molecular changes be, the word "mutation" typically refers to a change that affects the nucleic acids. In cellular organisms, these nucleic acids are the edifice blocks of Dna, and in viruses they are the building blocks of either DNA or RNA. One style to think of DNA and RNA is that they are substances that carry the long-term memory of the data required for an organism's reproduction. This article focuses on mutations in DNA, although we should keep in listen that RNA is bailiwick to essentially the same mutation forces.
If mutations occur in non-germline cells, then these changes tin can be categorized as somatic mutations. The give-and-take somatic comes from the Greek word soma which ways "trunk", and somatic mutations merely affect the present organism's trunk. From an evolutionary perspective, somatic mutations are uninteresting, unless they occur systematically and change some cardinal holding of an individual--such as the capacity for survival. For case, cancer is a stiff somatic mutation that volition affect a single organism's survival. Every bit a dissimilar focus, evolutionary theory is mostly interested in Deoxyribonucleic acid changes in the cells that produce the next generation.
Are Mutations Random?
The argument that mutations are random is both profoundly true and greatly untrue at the aforementioned time. The true aspect of this statement stems from the fact that, to the best of our cognition, the consequences of a mutation accept no influence whatever on the probability that this mutation volition or will not occur. In other words, mutations occur randomly with respect to whether their effects are useful. Thus, beneficial DNA changes practice non happen more often simply because an organism could benefit from them. Moreover, fifty-fifty if an organism has caused a beneficial mutation during its lifetime, the corresponding data volition not flow dorsum into the DNA in the organism'due south germline. This is a fundamental insight that Jean-Baptiste Lamarck got wrong and Charles Darwin got right.
However, the thought that mutations are random can be regarded as untrue if one considers the fact that not all types of mutations occur with equal probability. Rather, some occur more frequently than others considering they are favored by low-level biochemical reactions. These reactions are also the main reason why mutations are an inescapable property of any organization that is capable of reproduction in the real earth. Mutation rates are usually very low, and biological systems go to extraordinary lengths to keep them as low equally possible, by and large because many mutational effects are harmful. Nonetheless, mutation rates never reach zero, even despite both low-level protective mechanisms, like Deoxyribonucleic acid repair or proofreading during Dna replication, and high-level mechanisms, similar melanin deposition in skin cells to reduce radiation impairment. Beyond a certain betoken, avoiding mutation merely becomes too plush to cells. Thus, mutation will always be present as a powerful force in evolution.
Types of Mutations
So, how do mutations occur? The answer to this question is closely linked to the molecular details of how both DNA and the unabridged genome are organized. The smallest mutations are betoken mutations, in which simply a unmarried base pair is changed into another base pair. Yet another type of mutation is the nonsynonymous mutation, in which an amino acid sequence is inverse. Such mutations lead to either the production of a different protein or the premature termination of a protein.
Equally opposed to nonsynonymous mutations, synonymous mutations practice not change an amino acid sequence, although they occur, by definition, only in sequences that lawmaking for amino acids. Synonymous mutations exist because many amino acids are encoded by multiple codons. Base pairs tin as well have diverse regulating properties if they are located in introns, intergenic regions, or even inside the coding sequence of genes. For some historic reasons, all of these groups are often subsumed with synonymous mutations under the characterization "silent" mutations. Depending on their role, such silent mutations can exist anything from truly silent to extraordinarily important, the latter implying that working sequences are kept constant by purifying selection. This is the nigh probable explanation for the existence of ultraconserved noncoding elements that have survived for more than than 100 meg years without substantial alter, as institute past comparison the genomes of several vertebrates (Sandelin et al., 2004).
Mutations may also take the class of insertions or deletions, which are together known equally indels. Indels tin have a wide variety of lengths. At the short end of the spectrum, indels of one or two base pairs within coding sequences have the greatest effect, because they will inevitably cause a frameshift (just the addition of one or more than three-base-pair codons will keep a protein approximately intact). At the intermediate level, indels can affect parts of a gene or whole groups of genes. At the largest level, whole chromosomes or even whole copies of the genome can be affected by insertions or deletions, although such mutations are unremarkably no longer subsumed under the label indel. At this high level, it is likewise possible to invert or translocate entire sections of a chromosome, and chromosomes can fifty-fifty fuse or break apart. If a large number of genes are lost as a consequence of one of these processes, then the consequences are normally very harmful. Of course, unlike genetic systems react differently to such events.
Finally, still other sources of mutations are the many different types of transposable elements, which are small entities of DNA that possess a machinery that permits them to movement around within the genome. Some of these elements re-create and paste themselves into new locations, while others utilize a cut-and-paste method. Such movements can disrupt existing gene functions (by insertion in the heart of another factor), activate dormant gene functions (by perfect excision from a gene that was switched off by an earlier insertion), or occasionally lead to the production of new genes (by pasting cloth from different genes together).
Effects of Mutations
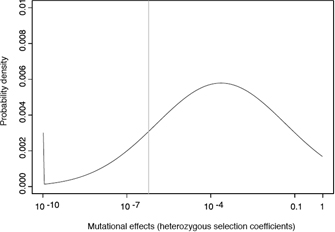
A single mutation can take a large upshot, but in many cases, evolutionary change is based on the aggregating of many mutations with small furnishings. Mutational furnishings can exist beneficial, harmful, or neutral, depending on their context or location. Most non-neutral mutations are deleterious. In full general, the more base of operations pairs that are affected by a mutation, the larger the effect of the mutation, and the larger the mutation'due south probability of existence deleterious.
To better understand the impact of mutations, researchers have started to estimate distributions of mutational effects (DMEs) that quantify how many mutations occur with what effect on a given property of a biological organisation. In evolutionary studies, the property of involvement is fettle, simply in molecular systems biology, other emerging properties might also be of interest. It is extraordinarily difficult to obtain reliable information about DMEs, because the respective effects span many orders of magnitude, from lethal to neutral to advantageous; in addition, many confounding factors usually complicate these analyses. To make things even more than difficult, many mutations also interact with each other to alter their effects; this phenomenon is referred to every bit epistasis. However, despite all these uncertainties, contempo piece of work has repeatedly indicated that the overwhelming majority of mutations have very small furnishings (Figure one; Eyre-Walker & Keightley, 2007). Of form, much more work is needed in order to obtain more detailed data about DMEs, which are a fundamental property that governs the development of every biological system.
Estimating Rates of Mutation
Many straight and indirect methods take been adult to help judge rates of dissimilar types of mutations in various organisms. The master difficulty in estimating rates of mutation involves the fact that DNA changes are extremely rare events and can merely be detected on a groundwork of identical Deoxyribonucleic acid. Because biological systems are usually influenced by many factors, direct estimates of mutation rates are desirable. Direct estimates typically involve apply of a known full-blooded in which all descendants inherited a well-divers Deoxyribonucleic acid sequence. To mensurate mutation rates using this method, i first needs to sequence many base pairs inside this region of DNA from many individuals in the pedigree, counting all the observed mutations. These observations are then combined with the number of generations that connect these individuals to compute the overall mutation rate (Haag-Liautard et al., 2007). Such directly estimates should non be confused with exchange rates estimated over phylogenetic time spans.
Summary
Mutation rates can vary within a genome and betwixt genomes. Much more work is required earlier researchers can obtain more precise estimates of the frequencies of different mutations. The rise of high-throughput genomic sequencing methods nurtures the hope that we will be able to cultivate a more detailed and precise understanding of mutation rates. Considering mutation is one of the cardinal forces of development, such piece of work will proceed to exist of paramount importance.
References and Recommended Reading
Drake, J. W., et al. Rates of spontaneous mutation. Genetics 148, 1667–1686 (1998)
Eyre-Walker, A., & Keightley, P. D. The distribution of fitness effects of new mutations. Nature Reviews Genetics 8, 610–618 (2007) doi:10.1038/nrg2146 (link to article)
Haag-Liautard, C., et al. Direct estimation of per nucleotide and genomic deleterious mutation rates in Drosophila. Nature 445, 82–85 (2007) doi:x.1038/nature05388 (link to article)
Loewe, L., & Charlesworth, B. Inferring the distribution of mutational effects on fitness in Drosophila. Biology Letters 2, 426–430 (2006)
Lynch, One thousand., et al. Perspective: Spontaneous deleterious mutation. Evolution 53, 645–663 (1999)
Orr, H. A. The genetic theory of adaptation: A brief history. Nature Review Genetics 6, 119–127 (2005) doi:10.1038/nrg1523 (link to commodity)
Sandelin, A., et al. Arrays of ultraconserved non-coding regions span the loci of key developmental genes in vertebrate genomes. BMC Genomics 5, 99 (2004)
godwinwerflefougs.blogspot.com
Source: http://www.nature.com/scitable/topicpage/genetic-mutation-1127
{kind=link}
Enregistrer un commentaire for "what does it mean to say that a mutation is deleterious?"